



מחלת הגרינינג היא כנראה מחלת ההדרים החמורה ביותר שידעה תעשיית ההדרים העולמית. המחלה נגרמת ע"י חיידק המותאם למחיה בצינורות השיפה ומועבר ע"י שני מיני פסילות הניזונות מהשיפה ומשמשות לו כווקטור. הפסילות נחשבות מזיק משני שאינו מזיק לעלי העץ, לפרחים ולפירות, אך עלולות לשאת את החיידק הגורם למחלת הגרינינג, לסייע בהפצתו ובכך לחרוץ את גורל התעשייה כולה.
מחלת הגרינינג
מחלת הגרינינג (greening), הידועה גם כ-Huanglongbing (HLB) (Yellow dragon disease, בסינית), כנראה התפתחה בצפון-מזרח סין על הדרים1. המחלה גורמת לכתמיות א-סימטרית בעלי ההדר, הצהבת ענפים, ירידה ביבול, הפרי קטן בעל אונות שונות בגודלן, מר בטעמו, חומצי מאוד ונושר בטרם עת. המחלה נקראת גרינינג בעקבות הצבע הירוק באזור פיטם הפרי לאחר ההבשלה2, בעוד שבתהליך הבשלה טבעי, ניכר צבע ירוק באזור העוקץ.
המחלה יכולה להיגרם ע"י אחד מ-3 טיפוסי חיידקים הנקראיםCandidatus Liberibacter africanus (CLaf), C. L. asiaticus (CLas), ו- C. L. americanus (CLam). החיידק האפריקאי (CLaf) נפוץ באפריקה3, רגיש לחום ופעיל בטמפ' של 22-24° (עד 27°)4. החיידק האסייתי (CLas) נפוץ באסיה ובאמריקה5, סביל/אוהב חום ופעיל בטמפ' חמות יותר של 32-35° (עד 38°)6. החיידק האמריקאי (CLam) מצוי בדרום אמריקה (ברזיל)7, סבילותו לחום נמוכה יותר והוא אינו פעיל מעל 32°6. החל משנת 2008, שכיחותו בפרדסים בברזיל ירדה משמעותית, בעוד זו של CLas עלתה (Ayres, A.J. pers. Comm.).
לחיידקים שני מיני וקטורים הנושאים אותם מצמח לצמח. פסילת ההדר האפריקאית Trioza erytreae (aka AfCP) (תמונה 1א) מצויה בעיקר באפריקה, אך גם באירופה (בספרד ופורטוגל)8, רגישה לטמפ' חמות ואינה מתפתחת בטמפ' מעל 30°9. פסילה זו מעבירה בעיקר את CLaf. פסילת ההדר האסייתית Diaphorina citri(aka ACP) (תמונה 1ב), נפוצה יותר ומצויה באסיה, אפריקה ואמריקה10. הפסילה מותאמת יותר לטמפ' החמות (25-28°) ואינה פעילה מעל 33°11. פסילה זו מעבירה בעיקר את CLas ו-CLam. ניכר שקיימת התאמה אקלימית בין החיידקים לפסילות, למרות ששתיהן יכולות לשאת את כל זני החיידקים12. שילוב של ACP על שלל תכונותיה עם אלו של CLas מהווים שילוב קטלני של מחלת הגרינינג בעולם.

פסילת ההדר האסייתית, ACP, נמצאה בישראל לראשונה בשנת 2021. סקרים להימצאות חיידק הגרינינג בארץ מבוצעים כל העת ע"י השירותים להגנת הצומח, משרד החקלאות. עד לכתיבת שורות אלו, לא נמצא אחד החיידקים בארץ.
פסילת ההדר האסייתית (ACP)
פסילות הן משפחה של חרקים Psyllidae השייכת לתת-סדרת הכנימות (Sternorrhyncha), יחד עם כנימות העש, כנימות העלה וכנימות המגן. לפסילת ההדר מחזור חיים בעל גלגול חסר, הכולל ביצים, נימפות ופסילה בוגרת ומשכו תלוי בתנאי הסביבה. הטמפ' האופטימלית לפסילה היא 25-28°, כאשר הפסילה משלימה מחזור חיים מביצה לבוגר ב-14-17 יום בממוצע. ככל שהטמפ' עולה, מחזור החיים מתקצר11.
הביצים מוארכות (0.3 מ"מ), צהבהבות ומוטלות על לבלוב צעיר, בקיפולי העלים לפני פתיחתם13, ובוקעות לאחר 3-4 ימים (תמונה 2א-ב). לאחר הבקיעה, הנימפה עוברת 5 דרגות התפתחות, כאשר עם ההתפתחות גודל גופן גדל, צבען הופך כהה יותר, עיניהן האדומות ניכרות וכן ניצני הכנפיים (תמונה 2ג-ה). בעוד הנימפות הצעירות ישיבות יותר, נימפות בדרגות 3-5 הן ניידות ומחפשות אתרי הזנה חדשים14. כל דרגות הנימפה ניזונות מהשיפה ובעקבות כך מפרישות גדילי שעווה אופייניים. בטמפ' אופטימלית, משך התפתחות הנימפות 11-13 יום11, ותלויה בזמינות לבלובים רכים להזנתם.
הפסילה הבוגרת קטנה יחסית (אורכה כ-4 מ"מ), גופה אפרפר עם פס חום בשולי הגוף ו'חלון לבן' בקצה (תמונה 1ב). הפסילות הבוגרות ניזונות מצינורות השיפה בחלקו התחתון של עלי הדרים צעירים ובוגרים. בעת ההזנה, ראשה קרוב לעלה, וגופה מורם למנח של 45° מפני העלה13. פעילותן מוגבלת לשעות היום במהלכו חלה ההזדווגות, הטלת הביצים והתעופה15. יכולת תעופתן אינו גבוה ומאופיין בעיקר בגיחות תעופה קצרות בתוך ובין פרדסים16. תעופה למרחק רב אפשרית בעזרת הרוח. הפרטים הבוגרים חיים כחודש, במהלכו הנקבה מזדווגת מס' פעמים בחייה עם זכרים שונים. יחס הזוויגים באוכלוסייה הוא 0.5 ללא תלות בצמח הפונדקאי17. הנקבה מטילה ביצים באופן רצוף לאורך חייה, כל עוד ישנם לבלובים זמינים, היא יכולה להטיל בין 620-750 ביצים בחייה11.
לפסילה 3 מופעים (morphotypes) של ACP: אפור/חום, כחול/ירוק וכתום/צהוב18. קיימים הבדלים בתכונות ביולוגיות ופונקציונאליות בין המופעים השונים19. לדוגמא, פסילות במופע הכתום/צהוב רגישות יותר לחומרי הדברה20. פסילות במופע הכחול/ירוק בעלות כשירות גבוהה יותר, יכולת תעופה טובה יותר ופוטנציאל רבייתי גבוה יותר מאשר שני המופעים האחרים18. בארץ אנו מזהים את המופע האפור/חום והמופע הכחול/ירוק (תמונה 2ו).

גודל אוכלוסיית הפסילה תלוי בזמינות לבלובים חדשים הדרושים להטלה והתפתחות הנימפות. עלייה משמעותית באוכלוסייה מתרחשת עם העלייה בזמינות הלבלובים בפרדס, בעונות הלבלוב העיקריות בסתיו ובאביב. נגיעות גבוהה של פסילות נמצאת לרוב בהיקף הפרדס מאשר בשורות הפנימיות21, ככל הנראה בגלל נוכחות גבוהה של לבלובים. הפסילה אינה מרבה לעוף למרחק רב (מקסימום 150 מ') וניכר שהיא מתיישבת בפונדקאי האופטימלי הראשון הנקרה בדרכה. לאחר אכלוס הפונדקאי, הפסילה עפה בצורה לא סדירה וכיוונית למרחקים נמוכים של כ-12-5 מטר22. מחקרים בודדים בספרות מדווחים על תעופה של כ-2 ק"מ במשך מס' ימים דרך בתי גידול שאינם מכילים פונדקאים מתאימים23,24.
לפסילות, גפי פה דוקרים-מוצצים (piercing-sucking mouthparts) באמצעותן הן מזריקות רוק לריכוך המזון באמצעות צינור אחד, ובאחר הן יונקות את המזון הנוזלי מצינור השיפה לגופן. עודף הסוכרים יוצא מגופה דרך פי הטבעת בצורת טל דבש. פסילות בוגרות מפרישות טיפה דביקה וצלולה הרחק מגופן, בעוד הנימפות מפרישות צינורות ארוכים של טל דבש עטופים בשכבה דקה של שעווה לבנה25. ההזנה על עלי הלבלוב גורמת לעיוות קצוות העלה בדומה לנזק הנובע מכנימות עלה26. בצפיפות אוכלוסייה גבוהה, הזנה של נימפות רבות מעלים מלבלבים תמנע את התפתחותם.
הפסילה ניזונה מצמחים ממשפחת הפיגמיים (Rutaceae). עיקר הפונדקאים בישראל הם הדרים (Citrus spp.), מוריאה מכבדית (Murraya paniculata) וקארי (M. koenigii). עצי הדר ובמיוחד שיחי מוריאה מכבדית נפוצים מאוד במרחב האורבני בישראל בגינות פרטיות, בבוסתנים וכצמחי נוי במרחב הציבורי. המוריאה המכבדית היא שיח רב שנתי המאופיין בגלי לבלוב רבים במהלך השנה, המהווים כר פורה להתפתחות אוכלוסיית הפסילה (תמונה 3). חוסר שימוש בתכשירי הדברה במרחב האורבני יחד עם שכיחות גבוהה של לבלובים המהווים אתרי הטלה רבים, מאפשרים התפתחות משמעותית של אוכלוסיית הפסילה ומקור (source) לפסילות הנצפות בפרדסים מסחריים כאשר אלו מלבלבים ומשמשים מבלע (sink) לאוכלוסייה זו. במחקר שבוצע בטקסס, פסילות מסומנות באבקה פלורסנטית צבעונית פוזרו בגבול בין שכונת מגורים לפרדס מסחרי, ונבחנה לכידת הפסילות בשני האזורים בשיטת MRR (mark-release recapture). כ-50% מהפסילות שפוזרו בשכונת המגורים, נלכדו מחדש בפרדס המסחרי הסמוך. לפיכך, המרחב האורבני אכן משמש אוכלוסיית מקור לנגיעות הפסילות בפרדסים הסמוכים27.

הפסילה והחיידק
דרך ההפצה העיקרי של המחלה הוא באמצעות הפסילות. החיידקים מוגבלים ומותאמים למחיה בצינורות השיפה בצמח28. הפסילה רוכשת את החיידק מהצמח בעת הזנתה מצינורות השיפה (בשלב הנימפה או הבוגר). רכישת החיידק יכולה לקחת 24 שעות, אך תלוי במשך ההזנה וברמת החיידק בצמח29. החיידק מתפשט בגוף הפסילה באמצעות ההמולימפה ומתרבה במעי. לאחר תקופת הדגרה של כ-10-15 יום30 בה אוכלוסיית החיידק עולה, מתחילה הפסילה להעביר את החיידק לצמחים חדשים מהם היא ניזונה. לפחות שעה של הזנה על הצמח החדש דרושה בכדי שיידבק בחיידק29. תחילה – אילוח הצמח מקומי, אך עם הזמן, אוכלוסיית החיידק בצמח גדלה, החיידק מתפשט בזרם השיפה לשורשים ולאזורים אחרים בנוף ופסילות נוספות יכולות לרכוש את החיידק ולהדביק עצים נוספים. הסימפטומים של המחלה יכולים להופיע לאחר מס' חודשים או רק כעבור שנתיים, מה שמקשה על מציאת העצים החולים בשלבים הראשונים של המחלה1. בכדי לוודא שמדובר במחלת הגרינינג, יש לבצע בדיקות מולקולריות, PCR, להימצאות ה DNA של החיידק בעלים חשודים או בפסילות.
פחות שכיחים הם מקרי הורשה אנכית (transovarial) של החיידק מנקבה לצאצאיה29 ובהעברה מינית במהלך הרבייה31. בעקבות כך, בצפיפויות גבוהות של אוכלוסיית הפסילה, החיידק יכול להישאר חיוני גם בהעדר מקור צמחי32.
הפצת המחלה ע"י הפסילה תלויה בגדילת אוכלוסייתה וביכולת פיזורה במרחב. מרכיב נוסף המסייע בהפצת המחלה נתון בכך שפסילות נאיביות נמשכות לעצים מאולחים בחיידק, ופסילות נגועות בחיידק נמשכות לעצים נקיים, מה שמעודד את התפשטות החיידק במרחב31. מודל סימולציה הראה שנדרשות רק 600 פסילות נגועות בחיידק במרכז הפרדס, בכדי להדביק את כל הפרדס במחלה תוך פחות משנה30. אך, כאשר הדבירו 75-90% מאוכלוסיית הפסילה, התפשטות החיידק בפרדס נבלמה לפחות בשנה או יותר. מסיבה זו, הדברה של וקטור המחלה, הפסילה, הינה קריטית להפחתת התפשטות החיידק לפרדסים נקיים והארכת פוריות העצים בפרדסים נגועים. חשוב לציין שאנו במדינת ישראל עדיין לא במצב זה. החיידק עדיין לא הגיע לישראל, אך כדי שנוכל להתמודד איתו טוב יותר לכשיגיע לארץ, והוא יגיע, כדאי מאוד שאוכלוסיית הפסילה בארץ תהיה נמוכה ככל האפשר.
השפעת האדם
בנוסף להפצת חיידקי הגרינינג ע"י הפסילה, מעבר בין צמחים יכול להתרחש גם דרך הרכבות. במקרה כזה, מעבר החיידקים תלוי בכמות חומר הריבוי וזן ההדר. לדוגמא, הרכבות עיניים נמצאו פחות יעילות למעבר החיידקים לעומת הרכבת רכב (side grafts of twigs)1. האיחוי העדין ברקמות הקמביום יוצר מעבר אופקי בין הכנה לרוכב המאפשר מעבר פתוגנים כדוגמת חיידקי הגרינינג. מעבר של CLas באמצעות הרכבה הוא פשוט, מהיר ויעיל יותר מאשר באמצעות הפסילה33, ומשמש כשיטה העיקרית להדבקת עצים למחקר מחלת הגרינינג.
שתילי מוריאה מכבדית והדרים מאולחים בפסילות, בחיידק או בפסילות הנושאות את החיידק עלולים להימכר במשתלות נוי ומרכזי גינון וכך להתפזר במרחב. ACP התפשטה בפלורידה כתוצאה ממשלוחי מוריאה מכבדית ממשתלות בדרום מחוז מיאמי, אשר הופצו דרך רשתות גינון34. בנוסף, תנועת חומר צמחי מאולח בפסילה (שתילים, פרי קטוף עם עלים ועוד), ללא ספק תרמה לפיזור המהיר של הפסילה בפלורידה35.
פלישת הפסילה או החיידק למדינה יכול להתרחש באמצעות מעבר לא מבוקר של פסילות בתעופה בגבולות יבשתיים בין מדינות. אולם, אפשרות נוספת היא פלישה באמצעות משלוחי חבילות מעבר לים. על פי ה USDA, נתפסו 170 משלוחים של ACP בעודה חיה ממדינות אסיה שונות בנמלים בארה"ב בשנים 1985-2003. בנוסף, נתפסו 73 משלוחים של צמחי מוריאה (לרוב קארי, אך גם הדרים) שעליהם 1ACP.
פסילת ההדר האסייתית, נמצאה לראשונה בישראל בעמק חפר ביולי 2021. בפרויקט משותף של השירותים להגנת הצומח (משרד החקלאות), ארגון המגדלים וענף ההדרים (מועצת הצמחים) ובשיתוף פעולה מלא מצד המגדלים, בוצעו פעולות הדברה מאסיביות שהביאו להיעלמות הפסילה מהאזור תוך 5 חודשים. שנה לאחר מכן, באוקטובר 2022 הפסילה נמצאה בעמק הירדן, ובעקבות מאמץ ניטורים נרחב, אותרה גם באזורים נוספים, למרות מאמצי ההדברה שננקטו. על-פי הניסיון של מדינות אחרות, מס' שנים לאחר פלישת הפסילה, פולש גם החיידק ואיתו מתפרצת מחלת הגרינינג (4 שנים בקליפורניה, 7 שנים בפלורידה). לפי המידע הקיים5, החיידק CLas הקרוב אלינו ביותר יבשתית נמצא בערב הסעודית, וייתכן שאף קיים במדינות השכנות לנו (אין לדעת). לא מן הנמנע שגם אלינו נכנסות פסילות לשטח המדינה בתעופה ממדינות שכנות ולא ניתן לשלול הגעת משלוחים מאולחים בפסילה ובחיידק מאסיה. ייתכן מאוד שמחלת הגרינינג, ואיתה האיום הקיומי על ענף ההדרים, כבר ממש בפתח.
מחלת הגרינינג – מקרי בוחן מהעולם*
*מתוך מאמר סקירה (Graham et al. 2024 review) 36
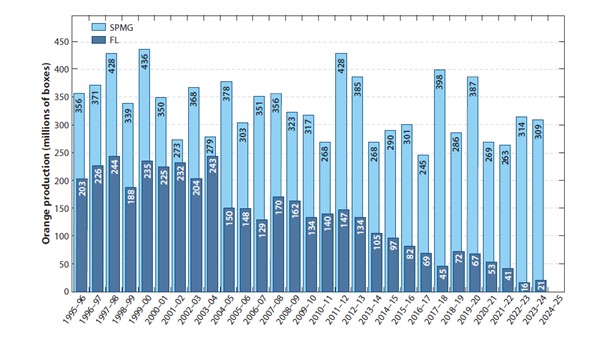
מדינות São Paulo (ברזיל) ופלורידה (ארה"ב) היו 2 אזורי ייצור תפוזים העיקריים בעולם, עד גילוי מחלת הגרינינג ב-2004 בברזיל וב-2005 בפלורידה. בשתי המדינות נקטו בניהול המצב בשיטת 3 הקודקודים: (1) נטיעה של שתילי הדרים נקיים מהמחלה, (2) הדברה יעילה של ACP ו-(3) עקירת עצים סימפטומטיים למחלה (תמונה 4). בברזיל, גישה זו הוכיחה את עצמה והביאה למעט ירידה יחסית להיקף התעשייה. בפלורידה לעומת זאת, ההשפעה על תעשיית ההדרים הייתה הרסנית, עם ירידה בתוצרת השנתית של כ-80%36 (תמונה 5).
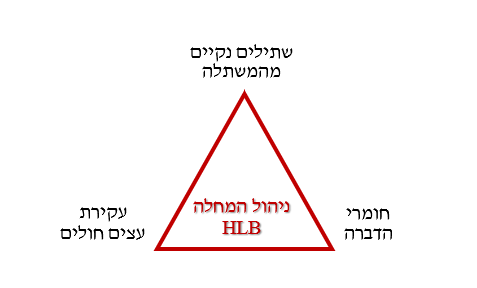
התוכנית התקבלה בצורה טובה ע"י המגדלים בברזיל, שכבר היו מנוסים בניטור עצים חולים, החלפתם בשתילים בריאים וטיפולי הדברה לווקטור המחלה, בעקבות מחלות קודמות כגון Citrus canker ו-CVC (Citrus variegated chlorosis). בשיתוף פעולה בין סוכנות הגנת הצומח של מדינת São Paulo וארגון Fundecitrus (ארגון פרטי הנתמך ע"י המגדלים ותעשיית המיץ), נקבעו רגולציות מנדטוריות להסרת עצים סימפטומטיים והחלפתם המיידית בעצים נקיים. למרות העדר אכיפה מחמירה, העקירה תרמה מאוד להאטה וצמצום התפשטות המחלה לאזורי הדרים אחרים בברזיל. עם השנים, הרגולציה התרחבה: פרדסים נטושים נעקרו או רוססו על ידי המגדלים מפרדסים סמוכים. הדרים ומוריאות במרחבים אורבניים ברדיוס של 5 ק"מ מפרדס מסחרי התבקשו להחליף צמחים אלו בצמחים אחרים שסיפק ארגון Fundecitrus (Miranda M, pers.comm.). הדברת הפסילה בחומרי הדברה הייתה בעייתית, עקב התפתחות עמידויות והגבלות שימוש לתוצרת המיוצאת ל-EU. כבר בהתחלה, חוקרי Fundecitrus הדגישו שטיפול בפסילה מחוץ לחלקה, לא פחות חשוב מטיפול בחלקה עצמה. ביוזמתם הוקם מערך של מגדלים מתנדבים שניטרו את הפסילות בחלקות הפרדס שלהם, לפיו בוצעו ריסוסים באזור נרחב בו זמנית (בתיאום בין החקלאים) בתוך ומחוץ לחלקה (כולל ריסוס פונדקאים לא נגועים) ע"י הפרדסנים.
בפלורידה, תוכנית שלושת הקודקודים אומצה ע"י ה- USDA, אך תחילה נדרשו כל משתלות ההדרים לייצר שתילים נקיים מ CLas בחממות סגורות שימנעו אילוח ע"י פסילות נגועות בחיידק. מחלת Citrus canker בפלורידה הביאה לעקירת מיליוני עצים מניבים וצעירים (1985-2005), ועתה גרמה להיענות נמוכה לעקירת העצים בקרב החקלאים. בעקבות כך, עקירת עצים סימפטומטיים התרחשה מס' מועט של שנים, עד שהוחלט שנגיעות המחלה נרחבת מדי, כך שהמשך העקירה לא ישאיר עצים כלל. הדברת הפסילה בחומרי הדברה בלבד, ללא עקירת עצים, לא הייתה מספקת למניעת התפשטות המחלה. עד 2013 החיידק התפשט כך שבכל פרדס היה עץ נגוע אחד לפחות. גם פרדסנים שריססו ועקרו עצים כנדרש, לא הצליחו לעצור את התפשטות המחלה. הפרדסנים התעלמו והייתה להם מעט השפעה על מקורות אילוח סמוכים לפרדסם, ולכן פסילות מאולחות בחיידק המשיכו להיכנס לפרדסם הנקי. בפלורידה, רוב החלקות קטנות וגובלות במס' פרדסים או עיירות, מה שמקשה על תיאום טיפול אזורי כפי שנעשה בברזיל, שם כל חווה מכילה שטחים נרחבים של הדרים.
בעוד שבפלורידה, חלק מהפרדסנים (בעיקר בסמוך לעיירות) מכרו את הקרקע ליזמים ובכך הורידו משמעותית את שטח הפרדסים במדינה, בברזיל, למרות העקירות הנרחבות, לא הייתה ירידה משמעותית בתוצרת בעקבות אופטימיזציה של ניהול הפרדס. זו כללה: התאמה טובה יותר של שילובי זן-כנה, שינוי מרחק הנטיעה (57 עצים לדונם במקום 39), השקיה מיטבית יותר (עלייה מ-5% ל-36% שטח מושקה) ודישון מתאים. כל אלו תרמו לעלייה ביבול לדונם. בחוות הטובות ביותר, היבול אף הוכפל לעומת היבול לפני הגעת מחלת הגרינינג.
כיום משתמשים בפלורידה במגוון טיפולים שונים להעלאת יבולים שיאפשרו לפרדסנים להמשיך לעבד את החלקות למס' שנים, כאשר הטיפול הבולט הוא הזרקת אנטיביוטיקה oxytetracycline לגזע. עם זאת, הפתרון שרוב המגדלים תולים בו את יהבם, היכול להציל את תעשיית ההדרים בפלורידה לטווח הארוך, הוא הנדסה (גנטית ולא גנטית) של עץ עמיד או סביל מספיק לחיידק. תהליך פיתוח זה יקר וארוך טווח ועצים אלו ייכנסו לשימוש רק אם תעשיית ההדרים בפלורידה תחזיק מעמד עד אז.
מהניסיון של מדינות אלו, אנו למדים שאסטרטגיה מאורגנת ומתואמת של ארגוני המגדלים, מכוני המחקר ובשיתוף פעולה מלא של החקלאים, כפי שנעשה בברזיל, היא האסטרטגיה המוצלחת יותר להתמודדות עם המחלה.
References
1Halbert, S.E. and Manjunath, K.L. 2004. Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: A literature review and assessment of risk in Florida. Florida Entomologist 83: 330-353.
2Bové, J.M. 2006. Huanglongbing: a destructive, newly emerging, century-old disease of citrus. Journal of plant pathology 88: 7-37.
3EPPO Global Database. 2024a. Candidatus Liberibacter africanus Distribution. Last update: 09.12.24. https://gd.eppo.int/taxon/LIBEAF/distribution
4Bové, J.M., Calavan, E.C., Capoor, S.P., Cortez, R.E. and Schwarz, R.E. 1974. Influence of temperature on symptoms of California stubborn, South Africa greening, India citrus decline, and Philippines leaf mottling diseases. Pages 12-15. In: Proceedings of the 6th Conference of the International Organization of Citrus Virologists, Swaziland.
5EPPO Global Database. 2024b. Candidatus Liberibacter asiaticus Distribution. Last update: 15.11.24. https://gd.eppo.int/taxon/LIBEAS/distribution
6Lopes, S.A., Frare, G.F., Bertolini, E., Cambra. M., Fernandes, N.G., Ayres, A.J., Marin, D.R. and Bové, J. M. 2009. Liberibacter associated with citrus huanglongbing in Brazil: ‘Candidatus Liberibacter asiaticus’ is heat tolerant, ‘Ca. L. americanus’ is heat sensitive. Plant Disease 93:257-262.
7EPPO Global Database. 2024c. Candidatus Liberibacter americanus Distribution. Last update: 15.11.24. https://gd.eppo.int/taxon/LIBEAM/distribution
8EPPO Global Database. 2024d. Trioza erytreae Distribution. Last update: 11.09.24. https://gd.eppo.int/taxon/TRIZER/distribution
9Pérez-Otero, R., Pérez-Turco, R., Neto, J. and Fereres, A. 2024. The African psyllid Trioza erytreae Del Guerio (1918) is very sensitive to low relative humidity and high temperatures. Insects 15: 62. doi: 10.3390/insects15010062
10EPPO Global Database. 2024e. Diaphorina citri Distribution. Last update: 12.12.24. https://gd.eppo.int/taxon/DIAACI/distribution
11Liu, Y.H. and Tsai, J.H. 2000. Effects of temperature on biology and life table parameters of the Asian citrus psyllid, Diaphorina citri Kuwayama (Homoptera: Psyllidae). Annals of applied biology 137: 201-206.
12Ajene, I.J., Khamis, F., Mohammed, S., Rasowo, B., Ombura, F.L., Pietersen, G., van Asch, B. and Ekesi, S.2019. First report of field population of Trioza erytreae carrying the Huanglongbing-associated pathogen, ‘Candidatus Liberibacter asiaticus’, in Ethiopia. Plant Disease Notes: 103: 1766.
13Hall, D.G., Richardson, M.L., Ammar, E.D. and Halbert, S.E. 2013. Asian citrus psyllid, Diaphorina citri, vector of citrus huanglongbing disease. Entomologia Experimentalis et Applicata. 146: 207-223.
14Tsai, J.H., and Liu Y.H. 2000. Biology of Diaphorina citri (Homoptera: Psyllidae) on four host plants. Journal of Economic Entomology 93: 1721-1725.
15Wenninger, E.J. and Hall, D.G. 2007. Daily timing of mating and age at reproductive maturity in Diaphorina citri (Hemiptera: Psyllidae). Florida Entomologist 90:715-722.
16Boina, D.R., Meyer, W.L., Onagbola, E.O. and Stelinski, L.L. 2009. Quantifying dispersal of Diaphorina citri (Hemiptera:Psyllidae) by immunomarking and potential impact of unmanaged groves on commercial citrus management. Environmental Entomology 38: 1250-1258.
17Nava, D.E., Torres, M.L.G., Rodrigues, M.D.L., Bento, J.M.S. and Parra, J.R.P. 2007. Biology of Diaphorina citri (Hem., Psyllidae) on different hosts and at different temperatures. Journal of Applied Entomology 131: 709-715.
18Wenninger, E.J., Stelinski, L.L. and Hall, D.G. 2009. Relationships between adult abdominal color and reproductive potential in Diaphorina citri (Hemiptera: Psyllidae). Annals of the Entomological Society of America. 102: 476-483.
19Martini, X., Hoyte, A. and Stelinski, L.L., 2014. Abdominal color of the Asian citrus psyllid (Hemiptera: Liviidae) is associated with flight capabilities. Annals of the Entomological Society of America 107: 842-847.
20Tiwari, S., Killiny, N., Mann, R.S., Wenninger, E.J. and Stelinski, L.L. 2013. Abdominal color of the Asian citrus psyllid, Diaphorina citri, is associated with susceptibility to various insecticides. Pest Management Science 69: 535-541.
21Gottwald, T. 2010. Current epidemiological understanding of citrus Huanglongbing. Annal Review of Phytopathology 48:119-139.
22Kobori, Y., Nakata, T., Ohto, Y. and Takasu, F. 2011. Dispersal of adult Asian citrus psyllid, Diaphorina citri Kuwayama (Homoptera: Psyllidae), the vector of citrus greening disease, in artificial release experiments. Applied Entomology and Zoology 46: 27–30.
23Martini X., Addison T., Fleming B., Jackson I., Pelz-Stelinski K. and Stelinski L.L. 2013. Occurrence of Diaphorina citri (Hemiptera: Liviidae) in an unexpected ecosystem: The Lake Kissimmee State Park Forest. Florida Entomologist 96: 658–660.
24Lewis-Rosenblum, H., Martini, X., Tiwari, S. and Stelinski, L. 2015. Seasonal movement patterns and long-range dispersal of Asian citrus psyllid in Florida citrus. Journal of Economic Entomology 108:3-10. doi:10.1093/jee/tou008
25Ammar, L.D., Alessandro, R., Shatters, R.G. Jr. and Hall, D.G. 2013. Behavioral, ultrastructural and chemical studies on the honeydew and waxy secretions by nymphs and adults of the Asian citrus psyllid Diaphorina citri (Hemiptera: Psyllidae). PLoS One 8: e64938. doi: 10.1371/journal.pone.0064938.
26Rogers, M.E. and Stansly, P.A. 2007. Biology and management of the Asian Citrus psyllid, Diaphorina citri Kuwayama, in Florida citrus. ENY-739. U of Florida, IFAS extension.
27Sétamou, M., Moreno, A.T. and Patt, J.M. 2022. Source or Sink? The role of residential host plants in Asian citrus psyllid infestation of commercial citrus groves. Journal of Economic Entomology 115. doi:10.1093/jee/toab249
28Thakuria, D., Chaliha, C.M., Dutta, P., Sinha, S., Uzir, P., Singh, S.B., Hazarika, S., Sahoo, L., Kharbikar, L.L. and Singh, D. 2023. Citrus Huanglongbing (HLB): Diagnostic and management options. Physiological and Molecular Plant Pathology 125:102016.
29Pelz-Stelinski, K.S., Brlansky, R.H., Ebert, T.A., and Rogers, M.E. 2010. Transmission parameters for Candidatus Liberibacter asiaticus by Asian citrus psyllid (Hemiptera: Psyllidae). Journal of Economic Entomology 103: 1531-1541.
30Lee, J.A., Halbert, S.E., Dawson, W.O., Robertson, C.J., Keesling, J.E. and Singer, B.H. 2015. Asymptomatic spread of huanglongbing and implications for disease control. Proceedings of the National Academy of Sciences 112: 7605-7610.
31Mann, R.S., Ali, J.G., Hermann, S.L., Tiwari, S., Pelz-Stelinski, K.S., Alborn, H.T. and Stelinski, L.L. 2012. Induced release of a plant-defence volatile ‘deceptively’ attracts insect vectors to plants infected with a bacterial pathogen. PLoS Pathogens. 8: p.e1002610.
32Grafton-Cardwell, E.E., Stelinski, L.L. and Stansly, P.A. 2013. Biology and management of Asian citrus psyllid, vector of the huanglongbing pathogens. Annual Review of Entomology 58: 413-432.
33Cui, X., Zhang, J., Liu, Y., Luo, X., Deng, X., Zhang, S. and Xu, M. 2022. Comparison of different grafting methods on the effect of ‘Candidatus Liberibacter asiaticus’ transmission. Fruit Research 2:15. doi.org/10.48130/FruRes-2022-0015
34Halbert, S., Manjunath, K.L., Ramadugu, C., Brodie, M.W., Wenn, S.E. and Lee, R.F. 2010. Trailers transporting oranges to processing plants move Asian citrus psyllids. Florida Entomologist 93: 33-38.
35Bayles, B.R., Shyam, T.M., Simmons, G.S., Grafton-Cardwell, E.E. and Daugherty, M.P. 2017. Spatiotemporal dynamics of the Southern California Asian citrus psyllid invasion. PLoS ONE 12: e0173226.
36Graham, J.H., Bassanezi, R.B., Dawson, W.O. and Dantzler, R. 2024. Management of Huanglongbing of citrus: Lessons from São Paulo and Florida. Annual Reviews of Phytopathology 62: 1-2.20. doi.org/10.1146/annurev-phyto-121423-041921






